
Citation
S. Pagliara, K. Franze, C.R. McClain, G.W. Wylde, C.L. Fisher, R.J.M. Franklin, A.J. Kabla, U.F. Keyser and K.J. Chalut
Nature Materials 13:638–644 (2014)
Abstract
Abstract
Embryonic stem cells (ESCs) self-renew in a state of naïve pluripotency in which they are competent to generate all somatic cells. It has been hypothesized that, before irreversibly committing, ESCs pass through at least one metastable transition state. This transition would represent a gateway for differentiation and reprogramming of somatic cells. Here, we show that during the transition, the nuclei of ESCs are auxetic: they exhibit a cross-sectional expansion when stretched and a cross-sectional contraction when compressed, and their stiffness increases under compression. We also show that the auxetic phenotype of transition ESC nuclei is driven at least in part by global chromatin decondensation. Through the regulation of molecular turnover in the differentiating nucleus by external forces, auxeticity could be a key element in mechanotransduction. Our findings highlight the importance of nuclear structure in the regulation of differentiation and reprogramming.
Figure sample
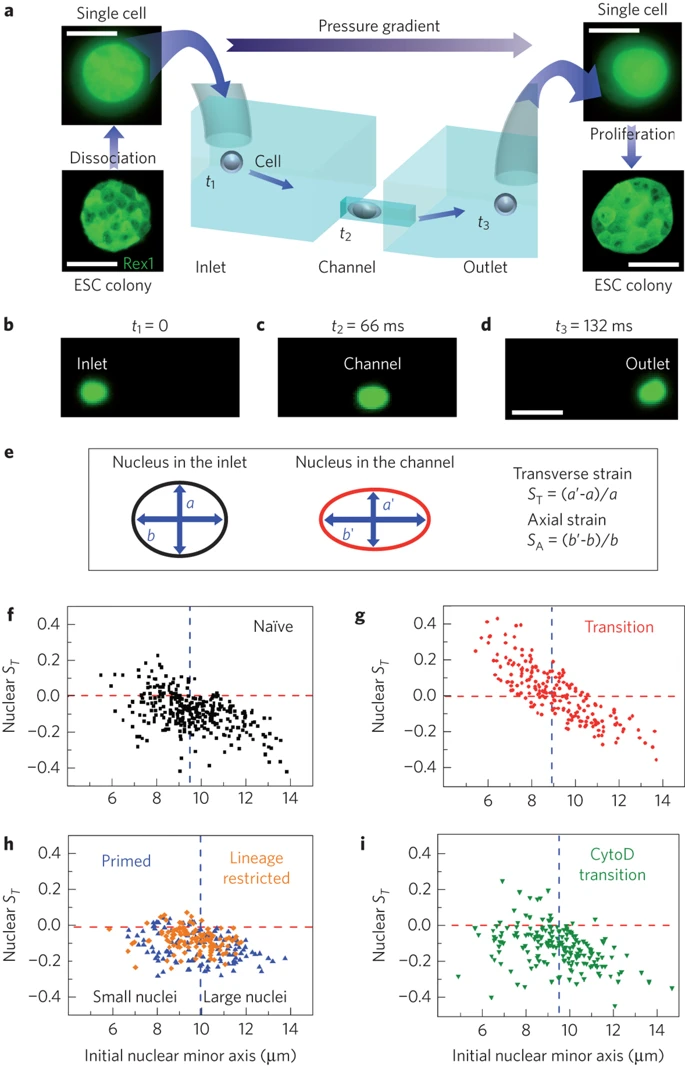
a, ESCs are detached from the culture plate, resuspended as single cells and loaded in the inlet of the microfluidic chip. The applied pressure gradient powers the transport of single cells from the inlet to the outlet. The inlet and outlet reservoirs are connected via channels smaller than the average cell size; therefore, cells must squeeze through the channels. Single cells collected from the outlet form colonies of fully functioning ESCs. Scale bars, 10 and 50 μm for single cells and colonies, respectively. b–d, Three consecutive epi-fluorescence snapshots of a Syto13-labelled cell translocating the channel from the inlet to the outlet under an applied pressure of 10 mbar. The detected fluorescence signal allows the simultaneous measurement of both cell and nuclear deformation. Scale bar, 25 μm. e, Schematic of nuclear shape whilst the cell is the inlet (left) and in the channel (right). We quantify nuclear deformation by using the transverse, ST, and axial strains, SA, where a (b) and a′ (b′) are the nuclear transverse (axial) extents in the inlet and in the channel, respectively. f–i, Scatter plots correlating the transverse nuclear strain ST in the channel to the initial nuclear minor axis for N-ESCs (squares), T-ESCs (circles), P-cells (upward triangles), lineage restricted HL60 cells (diamonds) and T-ESCs treated with Cytochalasin D (downward triangles). M = 3,3,2,1 and 2, respectively. A negative (positive) strain denotes a decrease (increase) of the nuclear transverse extent while the cell squeezes through the channel. The horizontal lines denotes a nuclear ST of zero and the vertical line is the population median for each data set used to separate the population of each class of cells in small and large nuclei categories. Small T-ESC nuclei show a markedly different behaviour from all other investigated cell types: their ST increases above 0, indicative of auxeticity.